|
FIG. 1. Diagram of the alligator
penis showing the morphometric indices used in this study. PL, length of the
penis tip; CD, diameter of the penis cuff. |
1Department of Zoology, 223 Bartram Hall, University of Florida, Gainesville, Florida 32611; and 2 National Biological Service, Florida Cooperative Fish and Wildlife Research Unit, Department of Wildlife Ecology and Conservation, University of Florida, Gainesville, Florida 32611
Accepted September 19, 1995
The development of the male reproductive ducts and external genitalia in vertebrates is dependent on elevated androgen concentrations during embryonic development and the period of postnatal growth. We have observed that a population of juvenile alligators living on Lake Apopka exhibit significantly smaller penis size (24% average decrease) and lower plasma concentrations of testosterone (70% lower concentrations) when compared to animals of similar size on Lake Woodruff. In addition to smaller phalli, no relationship exists between plasma testosterone concentrations and penile size in males from Lake Apopka, whereas a positive relationship exists for males from Lake Woodruff. The alligators on Lake Apopka are known to have elevated concentrations of the antiandrogenic DDT breakdown product p,p'-DDE stored in their fat. We suggest a number of hypotheses that could explain the modification in the phenotype of the juvenile male living in Lake Apopka. These modifications in phenotype include a smaller penis size, lower plasma androgen concentrations, and lack of responsiveness of the penis to the plasma androgens present.
In vertebrates, the development of the male internal reproductive ducts and external genitalia is largely dependent on the androgens testosterone and dihydrotestosterone. These sex steroids are responsible for the differentiation of the Wolffian duct into the male internal duct system as well as the formation of the external genitalia in reptiles (see Raynaud and Pieau, 1985). During embryonic development in alligators, secretion of androgens and Müllerian-inhibiting hormone from the testis produces an internal anatomy similar to that commonly described in other reptiles (Austin, 1989, 1990). In addition to embryologically organizing the male reproductive system, androgens are also responsible for stimulating the ontogenetic development of this system during juvenile (especially during puberty) and adult periods (Raynaud and Pieau, 1985). Additionally, seasonal fluctuations in the secretory activity and size of the testis and glands of the reproductive system are androgen dependent (Licht, 1984; Raynaud and Pieau, 1985).
Reptiles exhibit three distinctly different patterns of copulatory organ development, as the squamates (lizards and snakes) have paired hemipenes, the turtles and crocodiles (includes alligators and caiman) have a single penis, and the tuatara have no copulatory organ (Raynaud and Pieau, 1985). There has been an extensive debate on the homologies of the various copulatory organs in reptiles over the last century but a conclusion is still forthcoming. The available data suggest that the embryological development of the hemipenes in squamates is less complex than that seen in turtles (for a review see Raynaud and Pieau, 1985). Little is known of the embryological development of the copulatory organ in crocodilians. It is apparent that the crocodilian males and females have a homologous structure generally similar in gross morphology. It has been termed the cliteropenis by some workers (see Raynaud and Pieau, 1985; Allsteadt and Lang, 1995), whereas others use the term penis or phallus for this structure in males to denote its functional role (Forbes, 1939; Ramaswani and Jacob, 1965). Even in light of the possible lack of homology between squamate hemipenes and the chelonian or crocodilian penis, all available studies indicate that the steroid environment during embryonic development directly influences the development of the copulatory organ. That is, growth and differentiation of the penile anlagen in some reptilian species is stimulated by the presence of androgens, as reported for eutherian mammals, whereas in other species phallic development is inhibited by estrogens, as observed in birds (Raynaud and Pieau, 1985).
Androgens are essential for normal maturation and growth of the male reproductive system in reptiles. Seasonal growth of the testis produces elevated plasma concentrations of testosterone that are directly correlated with increased reproductive tract weight and synthesis of protein secretory products (Licht, 1984; Raynaud and Pieau, 1985). Exogenous androgen treatment of orchidectomized males reestablished reproductive tract growth and secretory activity. Previous studies have demonstrated that phallus growth is androgen dependent in alligators (Forbes, 1939) and crocodilians (Ramaswani and Jacob, 1965), as observed in other reptiles (Raynaud and Pieau, 1985) and in mammals (van Teinhoven, 1983). Interestingly, Ramaswani and Jacob (1965) observed that the penis of juvenile Indian mugger crocodile (Crocodylus palustris) exhibited the greatest androgen responsiveness of any male reproductive organ they examined, which included the secondary sexual ducts, the Wolffian duct, and the renal sex segment of the kidney. Likewise, recent studies have shown that significant penis growth in juvenile alligators can be induced by exogenous treatment with pharmacological levels of testosterone or dihydrotestosterone (DHT) (Jeff Lang, personal communication), whereas treatment with DHT at lower concentrations stimulated no significant growth in juvenile female alligators (Pickford, 1995). It should be noted that, in alligators, incubation temperature also influences the size of the clitoris in females, with females derived from eggs incubated at near male temperatures exhibiting larger clitorides than females from eggs incubated at lower temperatures (Allsteadt and Lang, 1995). However, the size, shape, and coloration of the phallic structures in 3month-old juvenile males and females are distinctive, allowing the correct diagnosis of sex 96% of the time (Allsteadt and Lang, 1995). Thus, phallus size could represent an obvious marker of abnormal androgen concentrations or functioning in reptiles as has been reported for mammals.
A number of environmental contaminants act as cellular signal transduction modifiers (Colborn et al., 1993; McLachlan, 1993; McLachlan et al., 1992). These contaminants interact with various cellular receptors-genetic transcription factors-so as to act as endocrine system agonists or antagonists. The vast majority of the compounds studied to date appear to bind to the vertebrate estrogen receptor, acting as xenoestrogens or antiestrogens (McLachlan, 1981, 1993; Safe et al., 1991; Soto et al., 1992, 1994; Peterson et al., 1993). Exposure to these compounds during embryonic development can have either catastrophic (e.g., mortality, cancer) or subtle (e.g., changes in enzyme function) effects as they are capable of modifying the organization of differentiating cells and organs (for discussion, see Guillette et al., 1995a).
A series of recent studies suggests that endocrine-disrupting contaminants can interact with other cellular receptor types, such as the androgen receptor (Gray et al., 1993; Kelce et al., 1994). For example, the fungicide vinclozolin is not an active endocrine system antagonist until it is metabolized to produce two compounds that act as antiandrogens. Exposure of developing rats to various doses of vinclozolin via maternal exposure produced a series of developmental abnormalities of the reproductive system in male offspring, including cryptorchidism, cleft phallus, hypospadias, and strophic seminal vesicles and prostate glands (Gray et al., 1993; Kelce et al., 1994).
Abnormalities of the male reproductive system have been reported in wildlife living in wetlands known to be contaminated with endocrine-disrupting chemicals (for a review see Colborn et al., 1993). For example, male alligators hatched from eggs collected from a contaminated lake in Florida, Lake Apopka, exhibit a number of developmental abnormalities such as depressed plasma testosterone concentrations, elevated testicular estradiol synthesis in vitro, and aberrant cellular structures in the seminiferous tubules (Guillette et al., 1994, 1995b). The underlying cause of these abnormalities, as well as the reported developmental problems in females, has been hypothesized to be due to "estrogenic" contaminants. The principal contaminant found in alligator eggs (Heinz et al., 1991) and juveniles (EPA, 1994) collected from Lake Apopka is p,p'-DDE, a major persistent metabolite of DDT. Although DDT has been reported to be estrogenic (Bulger and Kupfer, 1983; Fry and Toone, 1981), a controversy exists as to whether p,p'-DDE is estrogenic or not (see Soto et al., 1992, 1994; Kelce et al., 1995). However, using traditional receptor binding assays and a transfected yeast-androgen receptor system, Kelce et al. (1995) have demonstrated that p,p'-DDE binds the androgen receptor preferentially compared to the estrogen receptor and acts in the yeast system as an antiandrogen. It is the relative ratio of androgens to estrogens found in the plasma that creates an estrogenic or androgenic hormonal milieu. The presence of an antiandrogen would create an estrogenic environment by decreasing native androgen relative to estrogenic activity. These observations suggest the hypothesis that the abnormalities seen in the alligators from Lake Apopka may be due to an estrogenic environment produced not by the presence of an estrogenic contaminant but by the antiandrogenic action of p,p-'DDE. If elevated levels of an antiandrogenic compound were present in the tissues of embryonic and juvenile alligators at Lake Apopka, one would predict that androgen-dependent structures would be affected. Although p,p'-DDE is the primary suspect and is found in the greatest concentration in eggs and juveniles, it is important to note that many other compounds are present in Lake Apopka due to agricultural and municipal use and the concentrations of these compounds in eggs have not been examined. It is likely that the effects we observe in alligators could be due to the cumulative effect of a number of compounds. The purpose of this study is to examine the relationship between plasma testosterone concentrations and penis size in animals from a contaminated and a control lake. As penis size is dependent on circulating androgen concentrations during fetal and juvenile development, it represents an excellent marker of normal or abnormal steroid-induced development.
Animals and morphometrics. Juvenile alligators (Alligator mississippiensis) were collected at night by hand from two lakes in central Florida approximately 65 km apart. Lake Woodruff National Wildlife Refuge, Florida, was chosen as one study lake due to its relatively pristine nature as there is no agriculture or industry and only a few residences directly adjacent to the lake. On a single night (May 1994), 40 male and 15 female alligators were obtained from Lake Woodruff and adjacent marshlands. The following night, we hand-captured 54 males and 18 females from two localities on Lake Apopka, Florida. An additional 11 males and 2 females were caught on Lake Apopka during late April 1995. Sex was determined by the presence of a penis (dark pink to red in color) that could be extruded from the cloaca by manual pressure. Males averaged (±1 SEM, range) 35.7 cm (±0.7, 25.851.3) snout vent length (SVL), whereas females exhibited a mean SVL of 38.3 cm (±2.8, 23.0-63.0). Lake Apopka is a large eutrophic lake that has been exposed to various agricultural and municipal contaminants. Lake Apopka is also associated with a major pesticide (dicofol and DDT and its metabolites DDE and DDD) and sulfuric acid spill from the Tower Chemical Company that occurred in 1980 (U.S. EPA, unpublished report). Animals on Lake Apopka were captured from the Gourd Neck locality, adjacent to an unnamed stream that drained the Tower Chemical Company Superfund site. The Gourd Neck area was also extensively surrounded by citrus fields until the freezes of the mid-1980s. A second group of animals was collected along the northwest shore of the lake, approximately 8 km (linear shore length) from the Gourd Neck area.
Following capture, animals were placed in cloth bags and returned to a central site where a 1.5-ml blood sample was removed from the postcranial sinus of each animal. All samples were collected within 1 h of capture. Whole-blood samples were placed in a heparinized Vacutainer tube and stored on ice for 6 hr. Upon return to the laboratory, samples were centrifuged and plasma was collected, snap frozen in liquid nitrogen, and stored at -70° until assayed for plasma testosterone concentration (see assay description below).
After the blood sample was
collected, all animals were measured [total length (TL) and SVL] to the nearest
millimeter. The animals were then sexed by extrusion of the penis by manual
palpation of the base of the cloaca. Male and female alligators have a
cliterophallic structure that is similar in general shape although significantly
different in size, coloration (dark pink to dark red in males versus a very
light pink to white in females), and degree to which it can be manually extruded
from the cloaca (Allsteadt and Lang, 1995; L. J. Guillette, Jr., and A. R.
Woodward, unpublished data). If male, the length of the penis tip and width of
the penis cuff (see Fig. 1) were measured to the nearest 0.1 mm using a dial
caliper with needle tips. All measurements were made by a single researcher to
minimize and standardize measurement error. All animals were then tagged with a
unique toe tag and returned to their capture locality. For a detailed
description of gross genital morphology of the alligator, see Allsteadt and Lang
(1995) or Pickford (1995).
|
FIG. 1. Diagram of the alligator
penis showing the morphometric indices used in this study. PL, length of the
penis tip; CD, diameter of the penis cuff. |
Due to the greatly reduced size of the penis in many males from Lake Apopka, and the fact that the cliteropenis of male and female alligators is similar in basic structure (see Allsteadt and Lang, 1995; Pickford, 1995), a source of error in sexing animals is possible. That is, the greatly reduced male phallus could appear similar to a slightly enlarged female clitoris. Allsteadt and Lang (1995) have reported that some females from eggs incubated at elevated temperatures have an enlarged cliterophallus although it is always at the lower end of normal size in hatchling and young juveniles. However, the larger than normal cliterophallus in these females does not have the coloration of a male phallus nor can it be distended by manual pressure at the base of the cloaca as can be done with males. Nevertheless, we performed analyses on a subset of the data, in which all males having a phallus tip smaller than 3.0 mm were excluded. Juvenile females of the size used in this study do not have clitoris tips larger than approximately 2.0 mm and thus, if by chance we have included females in our "male" data set, the analysis of the subset would presumably remove this bias.
Radioimmunoassay (RIA). As significant seasonal variation can exist in plasma androgen concentrations, only the plasma from animals captured in May was assayed and reported under Results. Additionally, samples from 24 males were lost due to a freezer malfunction. Thus, the sample sizes are reduced compared to the available data for morphometric analyses.
Antisera for the androgen RIA were
obtained from Endocrine Sciences1 (Calabasas Hills, CA). The testosterone
antibody has the following cross-reactivities: dihydrotestosterone, 44%; ![]() -1-testosterone, 41%;
-1-testosterone, 41%;
![]() -1dihydrotestosterone, 18%;
5
-1dihydrotestosterone, 18%;
5![]() -androstan-30,17ß-diol, 3%;
4-androstan-3ß,17ß-diol, 2.5%; 5ß-androstan3ß,17ß-diol, 1.5%; estradiol,
0.5%; all other cross-reactivities are less than 0.2%. Radiolabeled testosterone
(TRK 402; 102 Ci/mmol) was purchased from Amersham Life Science, Inc. (Arlington
Heights, IL). Testosterone standard was purchased from Sigma Chemical Co. (St.
Louis, MO), and all other chemicals were obtained from Fisher Chemical Co.
(Pittsburgh, PA).
-androstan-30,17ß-diol, 3%;
4-androstan-3ß,17ß-diol, 2.5%; 5ß-androstan3ß,17ß-diol, 1.5%; estradiol,
0.5%; all other cross-reactivities are less than 0.2%. Radiolabeled testosterone
(TRK 402; 102 Ci/mmol) was purchased from Amersham Life Science, Inc. (Arlington
Heights, IL). Testosterone standard was purchased from Sigma Chemical Co. (St.
Louis, MO), and all other chemicals were obtained from Fisher Chemical Co.
(Pittsburgh, PA).
1 Use of manufacturer's name here and elsewhere does not imply endorsement by any of the authors or their respective institutions and agencies.
Alligator plasma samples (100 µl; all samples and standards performed in duplicate) were extracted twice with ethyl ether to remove the lipophilic steroids. After drying the sample ether extracts with filtered, low-humidity air, the samples were resuspended with 100 w1 borate buffer (0.5 M, pH 8.0). Androgen antibody (100 t,1; final dilution 1:36,000), BSA/ borate buffer (100 µl; 0.5 M borate buffer, 7.5% BSA),
and radiolabeled testosterone (100 µl of 9000 cpm) were added and the tubes were vortexed and incubated overnight at 4°. Bound-free separation was accomplished by adding 500 A.1 of 5% charcoal/0.5% dextran and centrifuging the tubes for 30 min at 2000 g. The supernatant was decanted, diluted with scintillation cocktail, and counted on a Beckman LS 5801 scintillation counter. Concentrations were estimated by commercially available software (Beckman Instruments).
Extraction efficiency was determined by spiking 100 µl of pooled alligator plasma with tritiated testosterone (approx 10,000 cpm) and determining the amount of label in the ether extract. To validate the androgen RIA for alligator plasma, plasma dilutions and internal standards were performed on a pool of alligator plasma samples. For plasma dilutions, plasma from the pool was aliquanted in 3-, 6-, 12-, 25-, 50-, 75-, and 100-µl amounts. These volumes were supplemented with plasma stripped of steroids (10 ml plasma + 1 g charcoal, 12-hr incubation, centrifuge and collect supernatant) such that all samples had 100 t.1. For internal standards, steroid-free plasma (100 A1) was spiked with 6.25, 12.5, 25, 50, 100, 200, 400, and 800 pg of testosterone. Both plasma dilutions and internal standards were extracted and assayed as described previously. Parallelism among the plasma dilution curve, internal standard curve, and standard curve was tested for homogeneity of slopes using the SuperANOVA statistical software program (Abacus Concepts, Berkeley, CA).
The minimum detectable dose in the assay was 29 pg/ml. Interassay variability was 15.6%, and intraassay variability averaged 4.3%. Extraction efficiency averaged 97%. The standard curve, plasma dilution curve, and internal standard curve were parallel, exhibiting homogeneity of slopes (F = 2.43, P = 0.127).
Statistics. Data on the length of the penis tip, diameter of the penis cuff, and plasma testosterone concentration were analyzed for differences among lake localities using one-way analysis of covariance (ANCOVA), with body size (total length or snout vent length) as the independent variable. Values for these parameters were log transformed prior to analysis to achieve homogeneity of variance. However, the least-squares means reported in the text are back-transformed to real values. Where significant (P < 0.05) variation existed, Scheffé's F test was used for
post hoc testing of individual paired comparisons. These analyses were performed using the SuperANOVA software program (Abacus Concepts, 1989). Additionally, regression analyses were performed comparing penis morphometric characteristics versus body size measurements (snout vent length, total body length) or plasma testosterone concentration of individual animals (Stat-View II, Abacus Concepts, 1988).
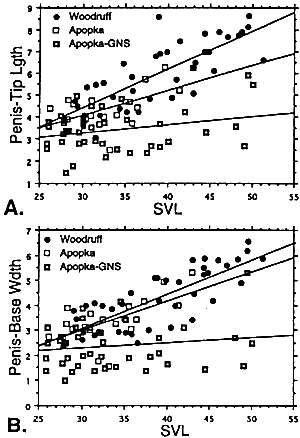
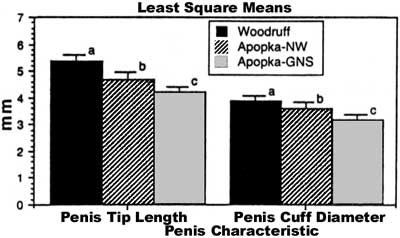
Penis morphometrics. We observed that the length of the penis tip and the diameter of the penis cuff were correlated with body size in juvenile males from two of three localities examined (Fig. 2). No relationship existed between penis size and body size in the Gourd Neck area of Lake Apopka (Fig. 2). Snout vent length explained over 59% of the variation in the diameter of the penis cuff and in the length of the penis tip on Lake Woodruff, whereas on the northwest shore of Lake Apopka, SVL was able to explain only 37% of the variation seen in the diameter of the penis cuff and 34% of the variation observed in the length of the penis tip. Although a relationship between body length and penis size, either tip length or cuff diameter, existed in two localities, a significant difference in the size of the penis was seen among all lake localities. Penis tip length (adjusted for TL, F = 3.8, df = 1,94, P = 0.025; adjusted for SVL, F = 3.5, df = 1,94, P = 0.03) and the diameter of the penis cuff (adjusted for TL, F = 3.8 df =1,94, P = 0.025; adjusted for SVL, F = 4.03, df =1,94, P = 0.02) were significantly larger on Lake Woodruff even after differences in body size were accounted for (Fig. 3). A significant difference in penis size was also observed between males from the two locations on Lake Apopka (Fig. 3). The difference observed in the length of the penis tip between animals caught on Lake Woodruff and Lake Apopka, after adjustment for snout vent length, indicated that a 24% difference in average length existed-13% reduction for animals caught on the northwest shore and 35% reduction for those caught in the Gourd Neck area. However, it is important to note that a difference in the length of the penis tip and the diameter of the penis cuff occurred in all size classes of juveniles when the two lakes were compared.
|
FIG. 2. Scattergram of the (A) length of the penis tip, or (B) diameter of the penis cuff versus snout vent length in three populations of juvenile alligators. Lake Apopka-NE: penis tip vs. SVL, r2 = 0.339, F = 9.2, P = 0.007; penis cuff vs. SVL, r2 = 0.371, F = 10.6, P = 0.004. Lake Apopka-GNS: penis tip vs SVL, r2 = 0.080, F = 3.4, P = 0.07; penis cuff vs SVL, r2 = 0.026, F =1.05, P = 0.31. Lake Woodruff: penis tip vs SVL, r2 = 0.591, F = 53.4, P < 0.0001; penis cuff vs SVL, r2 = 0.595, F = 54.3, P < 0.0001. |
|
FIG. 3. Least squares means (±1 SE) of the length of the penis tip and diameter of the penis cuff corrected for total body length in juvenile alligators from three lake localities in central Florida. Within a single morphometric parameter bars with different superscripts are significantly different. |
|
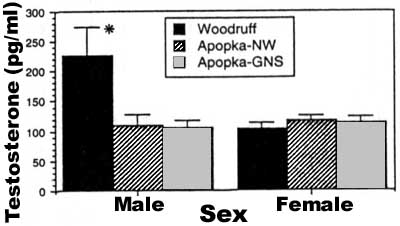
FIG. 4. Mean (±1 SE) plasma testosterone concentrations in male and female juvenile alligators from three lake localities in central Florida. *, significantly different from all other values. |
When all males having a penis tip length shorter than 3.0 mm are removed from the dataset (to remove the possible bias of females with enlarged clitorides sexed as males-see Materials and Methods), we observed that a significant difference still existed among all three populations following ANCOVA in penis tip length (adjusted for SVL, F = 6.18, df = 2,78, P = 0.003) and penis base diameter (adjusted for SVL, F = 5.1, df = 2,78, P = 0.008). A similar relationship existed when total body length was used as a covariate.
Plasma testosterone concentration. A comparison of plasma testosterone concentrations, corrected for body size, among lake localities and sexes indicated that males from Lake Woodruff had significantly higher concentrations (F = 4.9, df = 2,68, P = 0.01) than males found at either locality on Lake Apopka (Fig. 4). Males from both Apopka localities had plasma testosterone
concentrations similar to those exhibited by females from all three localities (Fig. 4). When plasma testosterone concentration was regressed with snout vent length, a significant correlation existed for males from Lake Woodruff (r2 = 0.212, F = 9.7, P = 0.004), whereas no correlation (P > 0.05) was observed for either locality on Lake Apopka (GNS area, r2 = 0.0003; NW area, r2 = 0.066). Likewise, no relationship between body size and plasma testosterone concentration was seen in females from any locality. The lack of a relationship between body size and plasma testosterone concentration is further clarified when mean concentration is determined for individual size classes of juvenile males. No apparent differences in plasma testosterone concentration existed for the smallest size classes when juvenile males from the two lakes were compared but dramatic differences existed for the two largest size classes where males from Lake Woodruff had 70% more plasma androgen than males of similar size from Lake Apopka (Fig. 5).
Finally, there was a positive relationship between penis tip length and plasma testosterone concentration in males from Lake Woodruff (F = 5.7, df = 1,36, P = 0.02), whereas we did not detect a relationship between these factors for males from either locality on Lake Apopka (GNS area, F = 0.02, df = 1,17, P = 0.9; NW area, F = 2.1, df = 1,18, P = 0.16). Penis cuff diameter exhibits a similar pattern with a positive relationship between plasma testosterone and cuff diameter for males from Lake Woodruff (F = 4.7, df = 1,36, P = 0.04) but no relationship was detected on Lake Apopka (GNS area, F = 0.08, df = 1,17, P = 0.8; NW area, F = 3.3, df = 1,18, P = 0.09).
|
FIG. 5. Percentage difference in mean lengths of the penis, diameters of the penis cuff, and plasma testosterone concentrations in different size classes of juvenile alligators obtained from Lake Apopka vs Lake Woodruff, Florida. Numbers above each size class represent the n for animals from Apopka /Woodruff. |
Male alligators from Lake Apopka had a significantly smaller phallus and lower plasma testosterone concentrations. Further, unlike males on the control lake, no relationship was observed between phallus size and plasma androgen concentration nor did phallus dimensions exhibit allometric relationships with body size for males obtained in the Gourd Neck area of Lake Apopka. We noted differences between collection sites on Lake Apopka. Juveniles living in the Gourd Neck area, near the entry site of the Tower Chemical Company pesticide spill, exhibited significantly reduced penis size and plasma testosterone when compared to juvenile male alligators living along the northwestern shore, which themselves exhibited reductions compared to the control lake. As reported previously for yearling alligators from Lake Apopka (Guillette et al., 1994), wild juvenile males collected during this study had plasma testosterone concentrations similar to those found in females.
The increased frequency of developmental abnormalities of male phallic structures, as reported in humans (Sharpe and Skakkebaek, 1993), fish (Davis and Bortone, 1992), and alligators (this study), is most easily explained by hypothesizing that androgen functioning has been reduced during embryonic development and early life. Several hypotheses can be developed to explain the reduced penis size in the juvenile male alligators from Lake Apopka: (1) males may have reduced plasma androgen concentrations that would result in reduced stimulation of the penis and, thus, reduced growth; (2) the relative ratio of testosterone to dihydrotestosterone could be altered by the suppression of the enzyme 5a-reductase, resulting in reduced penis size; (3) reduced numbers of androgen receptors on phallic tissue would also result in a reduction in size; (4) the presence of an androgen antagonist that would effectively compete for the androgen receptor and block normal androgen stimulation could also produce depressed phallus growth; and (5) the presence of a xenobiotic estrogen could shift the ratio of estrogens to androgens toward a feminizing environment in the developing embryo, thus blocking phallic development or inducing a female phenotype poorly responsive to androgens later in life. These hypotheses are not exclusive and all may contribute to a reduction in penis size to some degree.
The significantly reduced plasma testosterone concentrations could explain small phallus size in male alligators from Lake Apopka. Penis growth in male and female alligators and crocodiles can be stimulated by exogenous administration of either testosterone or dihydrotestosterone (Forbes, 1939; 1940; Ramaswani and Jacob, 1965; J. Lang, personal communication). Presumably, a lack or significant reduction in the circulating concentrations of these hormones would produce animals with reduced penis size. The data reported in this study confirm that sub-adult males of many age groups from Lake Apopka exhibit depressed plasma testosterone concentrations. We have previously shown that 6-month-old alligators hatched from eggs collected from Lake Apopka have severely reduced plasma concentrations of testosterone (Guillette et al., 1994). This study indicates that this reduction in plasma androgen is not transitory, occurring in neonates or yearlings only, but persists in animals much older in age.
What is responsible for the reduced plasma testosterone concentrations? When testes, obtained from juvenile alligators from a contaminated and a control lake, were cultured in vitro no difference in androgen synthesis was detected (Guillette et al., 1995b). We have hypothesized that the reduced plasma androgen levels in hatchling and juvenile alligators from Lake Apopka (Guillette et al., 1994) are due to modifications in liver degradation of androgens or modifications in the synthesis of plasma proteins responsible for binding steroids. (Guillette and Crain, 1995; Guillette et al., 1995b). Although mean plasma androgen concentrations are different between the males caught on the two lakes, it is very obvious that the major differences in plasma testosterone exist in the larger size classes. If animals are examined based on size class, it appears that males larger than 80 cm total length, from a pristine environment, show a significant increase in plasma testosterone concentration. These data suggest that plasma androgen concentrations do not exhibit a simple linear relationship with body size in juvenile alligators of all sizes. Moreover, since plasma androgen concentrations do not show elevations in larger sized males from Lake Apopka, we may conclude that the differences in penis size are exclusively due to the significant depression of plasma androgen alone. However, differences in penis size are also seen in very small alligators that had similar plasma androgen concentrations, suggesting that other factors may be involved with the observed differences. These factors appear to be acting during development or the early neonatal growth period.
A deficiency in the activity of the
enzyme 5![]() -reductase, responsible for the conversion of testosterone to
dihydrotestosterone, has been shown to produce a specific form of male
pseudohermaphroditism in which the external genitalia are poorly developed and
small in size (Impert-McGinley et al., 1979). Specifically, these males
have ambiguous or very small copulatory organs at birth that will, in some
individuals, grow with the onset of puberty. This pattern of abnormal
development is observed due to the specificity of androgen action during
embryonic develop
-reductase, responsible for the conversion of testosterone to
dihydrotestosterone, has been shown to produce a specific form of male
pseudohermaphroditism in which the external genitalia are poorly developed and
small in size (Impert-McGinley et al., 1979). Specifically, these males
have ambiguous or very small copulatory organs at birth that will, in some
individuals, grow with the onset of puberty. This pattern of abnormal
development is observed due to the specificity of androgen action during
embryonic develop
ment and organization of the male
genital anlagen. That is, the differentiation of the external genitalia and
prostate gland in male mammals appears dependent on adequate concentrations of
dihydrotestosterone, whereas testosterone controls the development of the
internal urogenital tract composed of the vas deferens, epididymis, and seminal
vesicles. Little data exist on the ratio of testosterone to DHT during normal
development in male reptiles. In the adult males examined, plasma testosterone
concentrations appear always to be significantly elevated over plasma DHT
concentrations (Bourne, 1991). We have no information on the activity of the
enzyme 5![]() -reductase but a preliminary study has shown that circulating
concentrations of DHT in juvenile alligators are very low or nondetectable with
no apparent difference between males from Lake Woodruff and Lake Apopka
(Pickford, 1995).
-reductase but a preliminary study has shown that circulating
concentrations of DHT in juvenile alligators are very low or nondetectable with
no apparent difference between males from Lake Woodruff and Lake Apopka
(Pickford, 1995).
Two alternative hypotheses to explain reduced phallus size are associated with either a reduction in androgen receptor frequency and distribution or the presence of an androgen antagonist-an antiandrogen. In mammals, the lack of androgen receptors-testicular feminization or androgen insensitivity syndrome-produces a condition where the male phenotype does not develop and XY individuals develop the external genitalia normal of females (Wilson and Foster, 1985). In mice, perinatal exposure to the synthetic estrogen diethylstilbestrol significantly reduces the number of androgen receptors in the prostate and seminal vesicle of male offspring (see Bern, 1992). Many of these mice exhibit reduced phallus size and modifications of the male reproductive tract. To our knowledge, there are no data on the frequency and distribution of androgen receptors in the alligator reproductive tract. Future studies must examine the natural distribution of androgen receptors in the reproductive system of embryonic, neonatal, juvenile, and sexually mature male and female reptiles.
Reduced androgen activity can be produced by the presence of an antiandrogenic substance. The effects of administration of antiandrogenic substances to developing mammals has been studied and one study examining the effect of antiandrogens during embryonic development has been performed in reptiles (Wibbels and Crews, 1992). Specifically, the administration of an antiandrogen, hydroxyflutamide, to embryonic red-eared turtles incubated at a temperature that induces 100% males, induced sex reversal-females-in 6% of the eggs treated (Wibbels and Crews, 1992). Neither the size nor the differentiation of the phallus was examined in this study (D. Crews, personal communication).
A number of environmental contaminants bind to hormone receptors and block normal hormonal functions. The fungicide vinclozolin, for example, is an androgen antagonist. Two metabolites of vinclozolin bind to the androgen receptor and block its function, causing demasculinization (but not complete feminization) of male rats exposed to this contaminant's breakdown products in utero (Kelce et al., 1994). Likewise, recent studies have indicated that a persistent metabolite of DDT, p,p'-DDE, is a relatively potent antiandrogen (Kelce et al., 1995), especially given its elevated concentrations in many organisms due to its ability to bioaccumulate and biomagnify in individuals feeding at higher levels of the food chain (for examples, see Clark and Krynitsky, 1985; Heinz et al., 1991; Rattner et al., 1984; Roelke et al., 1991; Subramanian et al., 1987; Williams et al., 1988). Juvenile alligators from Lake Apopka have elevated concentrations of p,p'-DDE (range 1.6-8.5 ppm) in their fat tissue (EPA, 1994). Alligator eggs collected from nests on Lake Apopka during the mid-1980s also had elevated concentrations of p,p'-DDE (Heinz et al., 1991). It is likely that this contaminant could act as an androgen antagonist in embryonic and juvenile alligators. In embryonic males, it could modify the organization of the penis, thus disrupting its responsiveness to an androgen signal later in life. This hypothesis may explain the absence of a relationship between phallus size and plasma testosterone concentrations in juvenile males from Lake Apopka. That is, a modification in the endocrine signals associated with embryonic development would consequently alter the responsiveness of male reproductive tract tissue to the activational stimulus of androgen later in life (for a general discussion of these concepts as they relate to contaminants, see Guillette et al., 1995a). Thus, the males from Lake Apopka could exhibit a "female" phenotype and exhibit poor responsiveness to androgen stimulation.
A number of hypotheses suggest
conditions that would produce male offspring lacking a phallus or with a phallus
reduced in size. Following their initial studies with the antiandrogenic
fungicide vinclozolin, E. Gray, W. Kelce, and colleagues proposed that the abnormalities of the penis and male
reproductive system seen in the rodents they treated could be due to a DHT
deficiency or receptor antagonism. Their follow-up studies indicated that 5![]() -reductase activity was not altered in the male offspring exposed to the
antiandrogenic metabolites of vinclozolin (Gray et al., 1993; Kelce et
al., 1994). Instead, the developmental abnormalities observed appeared to be
due directly to the inhibition of gene transcription by a blockage of the
activity of the androgen receptor (Kelce et al., 1994, 1995). p, p'-DDE
binds to the androgen receptor in vitro and blocks androgen
receptor-induced gene transcription in a transfected yeast assay (Kelce et
al., 1995). As p, p'-DDE represents the organochlorine pesticide found, to
date, to be in the greatest concentration in the alligators from Lake Apopka, it
suggests that future studies must examine the ability of this DDT metabolite to
bind the reptilian androgen receptor. Further, we must begin to understand what
factors) alters the development of male alligator embryos from Lake Apopka so as
to produce neonates and juveniles with reduced plasma androgen concentrations.
It is clear from the hypotheses discussed above that a number of interacting
factors could produce reduced phallus size. Future studies must test these
hypothesis and also determine if phallus reduction represents a biomarker of
permanent abnormalities of the reproductive system in these reptiles.
-reductase activity was not altered in the male offspring exposed to the
antiandrogenic metabolites of vinclozolin (Gray et al., 1993; Kelce et
al., 1994). Instead, the developmental abnormalities observed appeared to be
due directly to the inhibition of gene transcription by a blockage of the
activity of the androgen receptor (Kelce et al., 1994, 1995). p, p'-DDE
binds to the androgen receptor in vitro and blocks androgen
receptor-induced gene transcription in a transfected yeast assay (Kelce et
al., 1995). As p, p'-DDE represents the organochlorine pesticide found, to
date, to be in the greatest concentration in the alligators from Lake Apopka, it
suggests that future studies must examine the ability of this DDT metabolite to
bind the reptilian androgen receptor. Further, we must begin to understand what
factors) alters the development of male alligator embryos from Lake Apopka so as
to produce neonates and juveniles with reduced plasma androgen concentrations.
It is clear from the hypotheses discussed above that a number of interacting
factors could produce reduced phallus size. Future studies must test these
hypothesis and also determine if phallus reduction represents a biomarker of
permanent abnormalities of the reproductive system in these reptiles.
The St. John's River Water Management District, U.S. Environmental Protection Agency, and National Biological Service through Cooperative Agreement 11-16-00091544 RWO 137 with the University of Florida and grants to L.J.G. from the Environmental Protection Agency (CR821437) and the National Institute of Environmental Health (PR471470) provided partial funding for this project. We thank Allan Woodward, Dennis David, and other personnel of the Florida Game and Freshwater Fish Commission for their long-term support of our projects; Ab Abercrombie, Ken Rice, Jose Montes de Oca, and Isaac Chandler for assistance with collecting the juvenile alligators used for this study; and St. John's Water Management personnel for help with collecting juveniles on Lake Apopka; A. Cree and A. Woodward for providing very valuable comments on the manuscript as did two anonymous reviewers; and Leon Rhodes, Refuge Manager, Lake Woodruff National Wildlife Refuge, for providing assistance and allowing us to work on the refuge.
Allsteadt, J., and Lang, J. W. (1995). Sexual dimorphism in the genital morphology of young American alligators, Alligator mississippiensis. Herpetologica, in press.
Austin, H. B. (1989). Müllerian-duct regression in the American alligator (Alligator mississippiensis): Its morphology and testicular induction. J. Exp. Zool. 251, 329338.
Austin, H. B. (1990). The effects of estradiol and testosterone on Müllerian-duct regression in the American alligator (Alligator mississippiensis). Gen. Comp. Endocrinol. 76, 461-472.
Bern, H. (1992). The fragile fetus. In "Chemically-Induced Alterations in Sexual and Functional Development: The Wildlife/ Human Connection" (T. Colborn and C. Clement, Eds.), Vol. XXI, pp. 9-15. Princeton Sci. Pub., Princeton, NJ.
Bourne, A. (1991). Androgens. In "Vertebrate Endocrinology: Fundamental and Biomedical Implications," ( Pang, Ed.), Vol. 4, Part B, pp. 115-147. Academic Press, New York.
Bulger, W. H., and Kupfer, D. (1983). Estrogenic action of DDT analogs. Am. J. Indust. Med. 4, 163-173.
Clark, D. R., and Krynitsky, A. J. (1985). DDE residues and artificial incubation of loggerhead sea turtle eggs. Bull. Environ. Contam. Toxicol. 34, 121-125.
Colborn, T., vom Saal, F. S., and Soto, A. M. (1993). Developmental effects of endocrinedisrupting chemicals in wildlife and humans. Environ. Health Perspect. 101, 378-384.
Davis, W. P., and Bortone, S. A. (1992). Effects of kraft mill effluent on the sexuality of fishes: An environmental early warning? In "Chemically-Induced Alterations in Sexual and Functional Development: The Wildlife/Human Connection" (T. Colborn and C. Clement, Eds.), Vol. XXI, pp. 113-127. Princeton Sci. Pub., Princeton, NJ.
Environmental Protection Agency, U.S.A. (1994). "Tower Chemical Company Superfund Site Biological Assessment March 1994." U.S. Environmental Protection Agency, Biological Assessment.
Forbes, T. R. (1939). Studies on the reproductive system of the alligator. V. The effects of injections of testosterone proprionate in immature alligators. Anat. Rec. 75, 51-57.
Forbes, T. R. (1940). Studies on the reproductive system of the alligator. IV. Observations on the development of the gonad, the adrenal cortex, and the Müllerian duct. Contrib. Embryol. 174,131-163.
Fry, D. M., and Toone, C. K. (1981). DDT-induced feminization of gull embryos. Science 213, 922-924.
Gray, L. E., Jr., Ostby, J. M., and Marshall, R. (1993). The fungicide vinclozolin inhibits morphological sex differentiation in the male rat. Biol. Reprod. Suppl. 48, 97.
Guillette, L. J., Jr. (1994). "Developmental Abnormalities of the Reproductive System of Alligators (Alligator mississippiensis) from Contaminated and Control Lakes in Florida." Hearing on: "Health Effects of Estrogenic Pesticides." Subcommittee on Health and the Environment, Congressman Henry A. Waxman, Chairman, Testimony presented to U.S. House of Representatives 103-87; U.S. Govt. Printing Office, Washington, DC.
Guillette, L. J., Jr., and Crain, D. A. (1995). Endocrine-disrupting contaminants and reproductive abnormalities in reptiles. Commun. Toxicol., in press.
Guillette, L. J., Jr., Crain, D. A., Rooney, A. A., and Pickford, D. B. (1995a). Organization versus activation: The role of endocrinedisrupting contaminants (EDCs) during embryonic development in wildlife. Environ. Health Perspect. 103(Suppl. 7), 157 -164.
Guillette, L. J., Jr., Gross, T. S., Gross, D., Rooney, A. A., and Percival, H. F. (1995b). Gonadal steroidogenesis in vitro from juvenile alligators obtained from contaminated and control lakes. Environ. Health Perspect. 103(Suppl. 4), 31-36.
Guillette, L. J., Jr., Gross, T. S., Masson, G. R., Matter, J. M., Percival, H. F., and Woodward, A. R. (1994). Developmental abnormalities of the gonad and abnormal sex hormone concentrations in juvenile alligators from contaminated and control lakes in Florida. Environ. Health Perspect. 102, 680-688.
Heinz, G. H., Percival, H. F., and Jennings, M. L. (1991). Contaminants in American alligator eggs from lakes Apopka, Griffin and Okeechobee, Florida. Environ. Monit. Assess. 16, 277-285.
Imperto-McGinley, J., Peterson, R. E., and Gautier, T. (1979). Androgens and the evolution of male-gender identity among male pseudohermaphrodities with 5
-reductase deficiency. N. Engl. /. Med. 300, 1233-1237.
Kelce, W. R., Monosson, E., Gamcsik, M. P., Laws, S. C., and Gray, L. E. J. (1994). Environmental hormone disruptors: Evidence that vinclozolin developmental toxicity is mediated by antiandrogenic metabolites. Toxicol. Appl. Pharmacol. 126, 276-285.
Kelce, W. R., Stone, C. R., Laws, S. C., Gray, L. E., Kemppainen, J. A., and Wilson, E. M. (1995). Persistent DDT metabolite p,p-'DDE is a potent androgen receptor antagonist. Nature 375, 581-585.
Licht, P. (1984). Reptiles. In "Marshall's Physiology of Reproduction. Reproductive Cycles of Vertebrates" (G. E. Lamming, Ed.), Vol. 1, pp. 206-282. Churchill, Edinburgh.
McLachlan, J. A. (1981). Rodent models for perinatal exposure to diethylstilbestrol and their relation to human disease in the male. In "Developmental Effects of Diethylstilbestrol (DES) in Pregnancy" (A. L. Herbst and H. A. Bern, Eds.), pp. 148-157. Thieme, New York.
McLachlan, J. A. (1993). Functional toxicology: A new approach to detect biologically active xenobiotics. Environ. Health Perspect. 101, 386-387.
McLachlan, J. A., Newbold, R. R., Teng, C. T., and Korach, K. S. (1992). Environmental estrogens: orphan receptors and genetic imprinting. In "Chemically-Induced Alterations in Sexual and Functional Development: The Wildlife/Human Connection" (T. Colborn and C., Clement, Eds.), Vol. XXI, pp. 107-112. Princeton Sci. Pub., Princeton, NJ.
Peterson, R. E., Theobald, H. M., and Kimmel, G. L. (1993). Developmental and reproductive toxicology of dioxins and related compounds: Cross-species comparisons. Crit. Rev. Toxicol. 23, 283-335.
Pickford, D. (1995). Endocrine regulation of clitero-penis development in the juvenile American alligator (Alligator mississippiensis). Master's Thesis, pp. 137. University of Florida, Gainesville.
Ramaswami, L. S., and Jacob, D. (1965). Effect of testosterone proprionate on the urinogenital organs of immature crocodile Crocodylus palustris Lesson. Experientia 21, 206-207.
Rattner, B. A., Eroschenko, V. P., Fox, G. A., Fry, D. M., and Gorsline, J. (1984). Avian endocrine responses to environmental pollutants. /. Exp. Zool. 232, 683-689.
Raynaud, A., and Pieau, C. (1985). Embryonic development of the genital system. In "Biology of the Reptilia" (C. Gans and F. Billett, Eds.), Vol. 15, Development B, pp. 149-300. Wiley, New York.
Roelke, M. E., Schultz, D. P., Facemire, C. F., Sundlof, S. F., and Royals, H. E. (1991). Mercury contamination in the free-ranging endangered Florida panther (Fells concolor coryi). Proc. Am. Assoc. Zoo Vet. 1991, 277-283.
Safe, S., Astroff, B., Harris, M., Zacharwski, T., Dickerson, R., Romkes, M., and Biegel, L. (1991). 2,3,7,8-Tetrachlorodibenzo-p-dioxin (TCDD) and related compounds as antioestrogens: Characterization and mechanism of action. Pharmacol. Toxicol. 69, 400-409.
Sharpe, R. M., and Skakkebaek, N. E. (1993). Are oestrogens involved in falling sperm counts and disorders of the male reproductive tract? Lancet 341, 1392-1395.
Soto, A. M., and Chung, K. L., and Sonnenschein, C. (1994). The pesticides endosulfan, toxaphene and dieldrin have estrogenic
effects on human estrogen-sensitive cells. Environ. Health Perspect. 102, 380-383.
Soto, A. M., Lin, T.-M., Justicia, H., Silvia, R. M., and Sonnenschein, C. (1992). An "in culture" bioassay to assess the estrogenicity of xenobiotics. In "Chemically-Induced Alterations in Sexual and Functional Development: The Wildlife/Human Connection" (T. Colborn and C. Clement, Eds.), Vol. XXI, pp. 295-309. Princeton Sci. Pub., Princeton, NJ.
Subramanian, A. N., Tanabe, S., Tatsukawa, R., Saito, S., and Miyazaki, N. (1987). Reduction in the testosterone levels by PCBs and DDE in Dall's porpoises of Northwestern North Pacific. Mar. Pollut. Bull. 18, 643-646.
van Teinhoven, A. (1983). Reproductive Physiology of Vertebrates. Cornell Univ. Press, Ithaca, NY.
Wibbels, T., and Crews, D. (1992). Specificity of steroid hormoneinduced sex determination in a turtle. /. Endocrinol. 133, 121-129.
Williams, D., LeBel, G., and junkins, E. (1988). Organohalogen residues in human adipose autopsy samples from six Ontario municipalities. J. Assoc. Anal. Chem. 71, 410-414.
Wilson, J. D., and Foster, D. W. (1985). Williams Textbook of Endocrinology. Saunders, Philadelphia.
|
If you have come to this page from an outside location click here to get back to mindfully.org |